
Abstract
Climate change is associated with altered oceanographic conditions that tend to shift the geographical distributions of fish. To assess the impact of climate change on fisheries, one must go beyond projections of catch potential and understand how fishers respond to moving target species. Many previous studies have explicitly or implicitly assumed that fishers follow fish that are displaced by climate warming. Here, we evaluate this assumption by analyzing a long-term, large-scale yet high-resolution dataset combined with a detailed oceanographic model. Our study case is the Atlantic cod (Gadhus morhua) fishery in Norway, one of the largest whitefish fisheries in the world, with little technological or judicial constraints on the potential spatial response of fishers. An oceanographic model is used to predict the areas that have been suitable for Atlantic cod spawning over the two last decades. We compare whether these areas overlap with actually observed fishing locations. While the areas that are suitable for spawning clearly predict how much fish are caught per trip, the suitability of an area does not predict how many vessels fish in a given area at a given point in time. In contrast, the number of vessels in the previous week and the previous year explain the current number of vessels in that area. Hence, future projections of climate change effects should account for the rich and nuanced behavioral responses of humans to project climate change effects on fisheries.
Similar content being viewed by others


Introduction
Climate change profoundly affects oceanographic conditions and, as a consequence, the expected distribution of many fish species in the future1,2,3,4,5. Because harvested fish is an integral part of the world food system and because millions of people depend on fish caught in the ocean6,7,8, it is essential to assess how fishers respond to “species on the move”9.
Studies that project the effects of climate change on fisheries often assume, implicitly or explicitly, that fishers can follow fish10,11,12. However, there may be considerable constraints on fishers’ ability to follow target species. Especially in developing countries, available and affordable technologies limit how far fishers can move13. But also in industrialized countries, economic, regulatory, and behavioral factors reduce the adaptation potential of fisheries14,15. A better understanding of how fishers react to shifting species distributions with climate change is crucial to design adequate adaptation and mitigation policies. Yet, surprisingly little is known about the actual reactions of fishers to shifting species distributions16,17. Here, we match twenty years of individually observed fisher behavior with model-based predictions of fish distribution at a fine spatio-temporal resolution to assess the question of to what extent fishers follow fish.
To analyse how climate change affects fisheries, Cheung et al.18 calculate the so-called mean temperature of observed catch (the average preferred temperature of the species that are caught, weighted by their overall harvest). Using data from 52 large marine ecosystems between 1970 and 2006, the authors show a significant increase in the mean temperature of observed catch in the last decades18. Besides this global evidence, there is by now ample case-study evidence from regional studies19,20,21, and even for lake systems22 that highlights how climate warming affects fisheries. Yet, this metric cannot (and is not meant to) disentangle fish behavior from fisher behavior. In contrast, there are a small number of studies that match annual survey data from Alaska, the US East-, and West Coast with catch and landings data to differentiate between fish behavior and fisher behavior15,23,24,25. These studies mainly indicate a sluggish reaction of fishers to changes in the geographical distribution of target fish. We add to this literature by analyzing highly resolved data form Norwegian fishery on the Northeast Arctic (NEA) stock of Atlantic cod (Gadhus morhua). Hence, we can capture within-season behavioral change, which is an important margin for fishers. Moreover, in contrast to annual survey data, our predictions of fish distribution are directly based on exogenous oceanographic conditions that are independent of the current stock status.
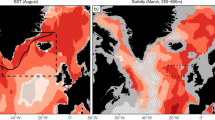
The Norwegian NEA cod fishery represents a significant share of global whitefish production. The annual first-hand sales revenue in this fishery is between 500 to 600 million Euro, placing it among the most valuable whitefish fishery in the world26,27. NEA cod is harvested with a variety of gears and boats, ranging from relatively small vessels that use hand-line and gillnets to large industrial trawlers. The main season of the NEA cod fishery is between mid-February and early May when the fish come to spawn close to shore along the coast of Northern Norway (Fig. 1).

The map shows the spatial resolution of the landings data (boxes) and 180 m depth contour (dotted line). The upper panel (inlet) shows the mean longitude of landings for the months Feb-May for each year (solid line) and the mean longitude of suitable habitat for the same months (dotted lines). The lower panel (inlet) shows the aggregated landings on a weekly basis for all years (solid black line) and for two example years: 2002 (Blue solid line) and 2018 (red solid line). For illustration, the area of the suitable spawning habitat (sum of all suitable grid points in the SVIM model; one model grid points represents an area of 16km2) is shown for all years (dotted black line) and for two example years: 2002 (blue dotted line) and 2018 (red dotted line).
These spawning aggregations make the NEA cod fishers an ideal case study because the oceanographic conditions are tightly associated with the suitability of an area for spawning28. Moreover, the relative proximity of the fishery to the Norwegian shore, and the broad variety of gear used to harvest NEA cod means that there are few technological or judicial constraints on the potential spatial response of fishers. In simple words: if the fish move with climate warming in our case study, and the fishers do not follow, it is not because they can’t, it is because they don’t want to. That said, our analysis does not aim to develop a structural model of fishers’ location choice in this specific fishery (see e.g.29,30,31). Rather, it is a first step that tests whether fishers follow fish that are displaced by climate warming. Our study thus opens the path for a more detailed investigation of fisher behavior under climate change in this and other cases.
Next to the socio-ecological setting of the study system, it is the availability of detailed data that allows us to answer our research question. The Norwegian Directorate of Fisheries publishes daily landing data per trip (2001–2021, 14.6 million observations). (The openly available data is anonymized, but vessel, gear, and further regulatory characteristics (e.g. which quota was used) are disclosed.) These so-called landing tickets contain, for each fish landing in Norway, information on species-specific weight and location of the catch, and by whom it was caught. We combine this data with output from an oceanographic model (see Methods and SI) to examine whether the number of vessels in a particular area at a given time can be explained by an index of spawning suitability for that area.
That is, we answer the question of whether fishers follow fish displaced by climate warming in three steps: First, we analyze how oceanographic conditions have changed in the recent past to establish whether the area suitable as NEA cod spawning sites has shifted. Then, we test the second central logical premise, namely whether oceanographic conditions indeed predict catch potential. Third, having these two pieces of evidence in hand, we present our main result: We document that fishers in one of the highest-value fisheries in the world have not followed fish over the past two decades.
Results
Changes in the oceanographic conditions suitable for spawning
Areas that are most suitable for spawning can change along two margins. On the one hand, areas that have historically been suitable may become suitable for spawning earlier in the season32,33. On the other hand, areas that have historically not been suitable for spawning may become suitable for spawning with climate change27,28.
We estimate spawning suitability for the years 2001–2021, following28: an area is suitable for spawning if the following three criteria were met: (i) local depth was less than 180 m, (ii) temperature was within the range of 4-6 degrees at some location between 50 and 150 m depth and (iii) salinity was within the range of 34.0–34.9 at some location between 50 and 150 m depth (“Methods”). We find that suitability changed along two margins: Suitable spawning conditions are observed earlier in the year, and more towards the north (Fig. 1, S2)34,35,36.
Moreover, Fig. 2 shows the weighted centroids of spawning suitability and the number of vessels operating cod fishing. The figure clearly shows that spawning suitability is more variable than observed harvesting locations. The suitability driven by natural conditions varies year by year while the fishers tend to stay in the same area and closer to shore.

Suitability for spawning predicts catch per trip
The second step in our investigation is to test whether fish actually follow suitable oceanographic conditions. The true abundance of fish in the water is not observed directly, but we can assess the catch potential by calculating catch per trip. To this end, we extract the quantity of harvested cod in a given trip of a given vessel from the landing tickets data. We then calculate the average catch per trip in a given area, aggregated to the weekly level to minimize the noise in the data and to reduce the effects of multi-day trips. We then test whether catch per trip in a given area in a given week is related to the spawning suitability of the area controlling for time, area, and vessel characteristics (gear and size).
Column (1) of Table 1 shows that the suitability explains the high catch per trip of cod. However, this sample contains many catch areas that are offshore and will permanently be unsuitable for spawning, e.g. because they are too deep or too far from the shore. Hence, we repeat the same regression on a restricted sample of those coastal areas that are north of 62∘N (column (2) in Table 1).
In both samples, we observed a positive and statistically significant effect of suitability on catch per trip. This result indicates that the model-based suitability index is a valid proxy for the abundance of cod stock at a detailed spatio-temporal scale. We controlled for various factors that could influence our results, such as fixed effects of time, space, and vessel attributes. In particular, we found that controlling for the vessel length group was crucial due to its strong effect on catch per trip. The time-invariant spatial effects are controlled by spatial fixed effects (area and region). The result demonstrates that the variation in suitability over time still explains catch in a given area even after controlling for the effect of ”always good” fishing grounds for fishers.
Suitability for spawning does not predict where vessels fish
Having established that (i) oceanographic conditions for spawning suitability change over time and (ii) that suitable habitats have more cod, we can now turn to the main question, namely whether the suitability for spawning of an area at a given time can explain the number of fishing vessels that operate in that area at that time. Results are presented in Table 2.
Column (1) of Table 2 shows the results from a regression model that explains the number of vessels in a given area at a given point in time by the suitability index, by the number of vessels in the same area a year ago, and by the number of vessels in the same area a week ago. The specification in column (2) additionally includes year fixed-effects to capture conditions that are specific to the year but the same for all vessels, such as the overall stock size, the annual quota, or general economic conditions, as well as week fixed-effects to capture the seasonality within a year. We see that the suitability of an area is a significant predictor of the number of vessels visiting that area in both these models.
However, once we control for area-specific effects in the specifications in columns (3) and (4), the suitability of an area no longer predicts the number of vessels. Interestingly, the number of vessels that were in a given area the week earlier, or a year earlier, remains a significant predictor for fishing effort, also when controlling for area-fixed effects.
These results are not consistent with the “fishers-follow-fish” hypothesis that postulates a positive coefficient on suitability. By comparing the specifications with varying sets of fixed effects, columns (3)–(4), we see that the inclusion of area-specific fixed effects absorbs the effect of suitability in both models. The suitability of an area is correlated with time-invariant area characteristics such as depth. The fishers know such characteristics based on their experiences. In addition, area fixed effects also control for economic reasons to visit a given area such as distances to home ports and the accessibility or perceived safety of the fishing ground.
An important question is how the link between the suitability for spawning and the number of vessels has developed over time. To study this question, we included an interaction between a linear time trend and the suitability index. We find suggestive evidence that the connection between the spawning suitability and the number of vessels in a given area is declining over time, but the effect is not statistically significant. While the literature finds the impact of climate change on NEA cod distribution with long-term data (e.g. 50 years), Fig. 3 illustrates these findings along with the model of annually varying coefficients.

The black line shows the estimates from a model with year dummies, and the dashed blue line indicates the estimate from a model with a linear trend. The error bars and the blue band show 95% confidence intervals.
Our results are robust to a variety of alternative models or sample specifications (see SI). The different selection of trip samples based on the share of cod in the total landing gives the same results (Table. S1–2). Including the neighborhood suitabilities also does not change the overall result (Table. S3). Moreover, using a recalculated suitability index to account for a potential modeling bias in the oceanographic model provides the result that the potential bias does not cause a large change in the result (Table. S4–5). The higher-order lag model was estimated to address potential mischaracterizations of the persistence in location choice, and the overall results remain consistent, as shown in Table S9. The analysis conducted with restricted samples, focusing on extreme years when suitability was concentrated either in the north-east or the south-west, suggests that fishers exhibit persistence in their location choices despite changes in suitability (Table S10).
Discussion
Fisheries are coupled socio-ecological systems that support the livelihood of millions of people around the globe. It is hence of key importance to assess how climate change will affect fisheries. To do so, one must both account for how the distribution of fish will change as fish follow their preferred oceanographic conditions, and assess how fishers react to the changes in fish distributions.
We find that spawning suitability can explain the number of vessels in the basic model, but once we control for area-specific effects (such as the distance to shore, or other features that do not shift through time), spawning suitability no longer explains the number of vessels in a given area. In contrast, the number of vessels in the same area the week or the year earlier continues to be a significant predictor of fishing effort. Therefore, our analysis does not support the hypothesis that fishers follow fish as fish following suitable oceanographic conditions, at least over the time horizon that we study in this paper. Rather, our results are in line with fishers exhibiting “site fidelity”, even in an industrialized fishery with little technological or regulatory constraints on mobility37,38.
That said, one has to keep in mind that our sample period may be too short to capture the impact of climate change, or it could be that the observed temperature change has been too small to affect habitat suitability to such an extent that it would also affect fishers’ harvesting locations. In Fig S4 and S5 of the SI, we show that there is both significant intra-annual and inter-annual variation in suitability. It is difficult to extrapolate fisher behavior to conditions when habitats shifts outside of the range of observed seasonal variation for prolonged periods. Our results suggest that the discrepancy between the fishers and the fish locations may become larger as climate change would affect the suitability more severely and the variations are expected to become greater. While the subdued variations in responsiveness suggest a limited adaptation capacity of the fishing sector to climate change, the situation may look different when conditions become more extreme.
An important early study that differentiates between fish behavior and fisher behavior is the work of Pinsky and Fogarty23. The authors contrast the average latitude at which a species occurs in annual research surveys with the average latitude at which the same species is landed in commercial catches in the same year (for the US Atlantic coast) and show that fishers do follow fish, but with a lag. Pinsky and Fogarty highlight that one of the reasons why fish landings shifted only by 10-30% could be that fishers increased harvesting pressure as the stock was shifting out of their historical habitat. Such a compensatory stock depletion would yield high harvest39 and mask the effect that would be visible if fishing effort were proportional to abundance. An important advantage of our study case is that the migratory and seasonal nature of the NEA cod fishery means that fishers harvest at capacity during the spawning season. Thus, there is no compensatory depletion effect, at least in the short run.
More generally, our estimate of the geographical distribution of the target fish is based on a physical model that is, by construction, independent of fishers harvesting behavior. We find that spawning cod are likely to be observed earlier in the year, and more towards the north. Our model predictions thus replicate earlier findings34,35,36.
Certainly, such model-based projections of fish distribution come with the important caveat that they are but an imperfect proxy of the true distribution, which is not observed. That is why the first step of our analysis, showing the significant relation between the spawning suitability of an area and the catch per trip in that area is important and reassuring.
A central feature of our study is the high spatial and temporal resolution of our data. This resolution allows us to assess intra-seasonal behavioral responses, which are an important margin for the profit maximization of fishing firms. We find that fishers did not follow fish as fish follows suitable oceanographic conditions in the NEA cod fishery between 2001 and 2021. Spawning suitability does not predict the number of vessels in a given area at a given point in time once we control for area-fixed effects. In contrast, the number of vessels in that area one week prior, or one year ago, remains a significant predictor of where fishers go.
For the interpretation of this result, it is important to appreciate that we do not structurally model fisher behavior. Instead, we conduct a “reduced form” estimation that can test our hypothesis but does not uncover the specific mechanisms at play. The area-fixed effect absorbs many important economic conditions that are constant for a given area, such as the distance to the nearest port (and hence travel cost) or the differential exposure to wind and waves. For example, the fact that the number of vessels that have historically been active in a given area is a significant predictor of current activity may reflect that fishers are constrained in the information they have available when forming expectations about which area to go to31. If the information on spawning suitability can be made available in real-time to fishers, this could be a business opportunity for private firms or a welfare-enhancing policy for public regulators.
Indeed, a better understanding of how fishers behaviorally respond to changes in species distribution is essential for effective and resilient resource management. Young et al.40, for example, analyzed adaption strategies in the US Atlantic coast. A key contribution of Young et al.40 is to study the location of fishing (as opposed to the location of landing) at a high resolution. Our approach allows us to do so as well, but in contrast to40, we focus on one species only and match the fishing location with the predicted fish location. In other words, our focus is not so much on fishers’ global adaptation strategies, but more on their spatio-temporal mobility as a response to shifts in the availability of their main target species.
In addition to mobility, another adaptation strategy is catch diversification41,42. Catch diversification not only allows fishers to buffer current risks43, but it may also prepare fishers to buffer future risks. Young et al.40, for example, show that vessels with historically lower catch diversity are less likely to persist over time compared to vessels with higher catch diversity. Yet, specialization versus diversification involves a considerable economic trade-off. Analyzing how such a trade-off plays out under expected climate change is an important avenue for future research.
A number of studies analyze the vulnerability of fishing communities to future climate change risks24,44,45. Our study complements this growing literature by matching actually observed fisher behavior with model-based fish behavior. It hence provides a ground truthing for such vulnerability assessments. We find that fishers do not seem to follow the fish, even in a technologically advanced fishery with little regulatory constraints on where fishers go.
For the specific case of the NEA cod fishery, this is good news, as it may reduce tensions between Russia and Norway that share this fishery. Game-theoretical work has highlighted how the strategic interactions between the two countries may be adversely affected by the shift in distribution when fishers follow fish46,47. (As one player, Russia in this case, gains more access to the stock and the other player, Norway in this case, loses access to the stock, competition intensifies, which may jeopardize the sustainability of the fishery39.) When fishers do not follow the shift in fish distribution (or at least to a lesser extent), this may lead to less competition than originally anticipated.
Whether the finding that fishers do not follow fish as fish follow suitable oceanographic conditions is good news more generally is an open question. Certainly, it remains important to anticipate and prepare for shifting geographical locations of target species due to climate change9. Our study emphasizes that future projections must account for the rich and nuanced behavioral responses of humans to assess climate change effects on fisheries.
Methods
Data sources
The source of the landing ticket data is the Directorate of Fisheries. (https://www.fiskeridir.no/Statistikk/AApne-data/AApne-datasett/Fangstdata-koblet-med-fartoeydata) The landing tickets specify, each time a vessel lands fish in Norway, which species are landed (landings are differentiated by typical market categories and mainly recorded in terms of weight, but sometimes also in terms of individual specimens that were caught). Additionally, the data contains information about the vessel that was used to catch the fish and the self-reported area where the fish was caught. The GPS coordinates of the catch areas (”fangstfelt”) is downloaded from the map (https://kart.fiskeridir.no/fiskeri), ”statistikkområder” then ”Lokasjoner (t.o.m. 2017)”. These areas are our basic unit of analysis and we count the number of vessels that fish in a given area in a given week in a given year. We aggregate the landing ticket data to the weekly level despite the fact that it is available with a daily resolution to minimize noise and to account for multi-day trips.
To estimate the suitability for spawning of a catch location ("fangstfelt”) at a certain day in a certain year, we used the output from an oceanographic model. The model used was the Regional Ocean Modeling System (ROMS,48,49) applied to the Northeast Atlantic Ocean (SVIM,50). A hindcast archive has been established for this model covering the period 1959 to the present. The spatial horizontal resolution of the model run was 4 × 4 km, which gives a grid cell count for each of the rectangles in Fig. 1 varying from 180 points in the south to 120 points in the north. The model has been validated and is considered to perform well51, especially with regard to modeled temperatures. However, salinity is less well represented in the model and we have correspondingly conducted sensitivity tests (SI).
For most of the analysis, we concentrate on coastal areas. The spawning sites of NEA cod are distributed along the coast and the main fleet targeting this aggregated stock is the coastal fleet. Coastal areas in this study are defined as the main statistical areas (Hovedområder) that include 12 nautical miles lines (Figure S1).
Suitability index
Following28, we defined that a grid point in the oceanographic model was suitable for spawning if the following three criteria were met:
-
1.
local depth was less than 180 m,
-
2.
Temperature within the range of 4–6 degrees at some location between 50 and 150 m depth
-
3.
Salinity within the range of 34.0–34.9 at some location between 50 and150 m depth.
The suitability was assessed daily for each grid point over the period with reliable data on landings at the catch area scale (2001–2021) between February and May, covering the spawning season of NEA cod. To facilitate comparison with the landing ticket data, the suitability was aggregated inside each catch area and over weeks. This was done by assessing the daily percentage of suitable grid points inside each catch area and then averaging over weeks. Note that we consider the fraction of suitable areas and not the fraction of suitable volume as cod tend to separate into two distinct depth groups with female distributed higher up in the water column compared to males52. Hence, we consider an area-based index more appropriate than a volume-based index.
Empirical methods
We model the total number of vessels fishing in an area, Niwya, for fleet i during week w in year y in area a as
where αj represents the fixed effects (varying intercept) for year, week, area (fangstfelt), region (hovedområde), and fleet. A fleet is defined as the combination of the gear that is used and the length group of the vessel. Swya is the suitability index.
We estimate model (1) using the R-package fixest, see53. The main test is whether the coefficient β0 is positive. If it is, then we can reject the null hypothesis that there is no relationship between the suitability of an area for spawning at a given point in time and the number of vessels in that area at the same point in time. In addition to testing this hypothesis for varying sets of fixed-effects αj, we include two additional controls in the regressions: Ni[w−1]ya, the number of vessels in the same area and the same year but a week earlier, and Niw[y−1]a, the number of vessels in the same area and the calendar week but a year earlier. Finally, we include an interaction of the suitability index and a linear time trend in one specification to test whether the association between the location of the fishing effort and the suitability is changing over time. This interaction term is not reflected in the regression model (1), but the result is shown in Fig. 3.
Data availability
The data that support the findings of this study are openly available at the Directorate of Fisheries at https://www.fiskeridir.no/English/Fisheries/Statisticsand https://www.fiskeridir.no/Tall-og-analyse/AApne-data/Fangstdata-seddel-koblet-med-fartoeydata. The results from the numerical model results are openly available. They were originally published on https://archive.norstore.no/pages/public/datasetDetail.jsf?id=10.11582/2015.0001454, but limited to a period ending in 2011. The simulation was subsequently extended in time, and the full period is openly available as a sub-settable dataset from https://thredds.met.no/thredds/nansen-legacy.html.
References
Pinsky, M. L., Worm, B., Fogarty, M. J., Sarmiento, J. L. & Levin, S. A. Marine taxa track local climate velocities. Science 341, 1239–1242 (2013).
Poloczanska, E. S. et al. Global imprint of climate change on marine life. Nat. Clim. Change 3, 919–925 (2013).
Booth, D. J., Feary, D., Kobayashi, D., Luiz, O. & Nakamura, Y. Tropical Marine Fishes and Fisheries and Climate Change, Ch. 26, 875–896 (John Wiley & Sons, Ltd, 2018).
Free, C. et al. Impacts of historical warming on marine fisheries production. Science 363, 979–983 (2019).
Gervais, C. R., Champion, C. & Pecl, G. T. Species on the move around the australian coastline: A continental-scale review of climate-driven species redistribution in marine systems. Global Change Biol. 27, 3200–3217 (2021).
Smith, M. D. et al. Sustainability and global seafood. Science 327, 784–786 (2010).
Hilborn, R. et al. Effective fisheries management instrumental in improving fish stock status. Proc. Natl. Acad. Sci. 117, 2218–2224 (2020).
Costello, C. et al. The future of food from the sea. Nature 588, 95–100 (2020).
Pinsky, M. L. et al. Preparing ocean governance for species on the move. Science 360, 1189–1191 (2018).
Cheung, W. et al. Large-scale redistribution of maximum fisheries catch potential in the global ocean under climate change. Global Change Biol. 16, 24–35 (2010).
Erauskin-Extramiana, M. et al. Large-scale distribution of tuna species in a warming ocean. Global Change Biol. 25, 2043–2060 (2019).
Tai, T. C., Steiner, N. S., Hoover, C., Cheung, W. W. & Sumaila, U. R. Evaluating present and future potential of arctic fisheries in canada. Marine Policy 108, 103637 (2019).
Belhabib, D., Lam, V. W. & Cheung, W. W. Overview of west african fisheries under climate change: Impacts, vulnerabilities and adaptive responses of the artisanal and industrial sectors. Marine Policy 71, 15–28 (2016).
Haynie, A. C. & Pfeiffer, L. Why economics matters for understanding the effects of climate change on fisheries. ICES J. Marine Sci. 69, 1160–1167 (2012).
Haynie, A. C. & Pfeiffer, L. Climatic and economic drivers of the bering sea walleye pollock (theragra chalcogramma) fishery: implications for the future. Can. J. Fish. Aquatic Sci. 70, 841–853 (2013).
Papaioannou, E. A. et al. Not all those who wander are lost - responses of fishers’ communities to shifts in the distribution and abundance of fish. Front. Marine Sci. 8, 669094 (2021).
Samhouri, J. F. et al. Stay or go? geographic variation in risks due to climate change for fishing fleets that adapt in-place or adapt on-the-move. PLOS Clim. 3, 1–28 (2024).
Cheung, W. W. L., Watson, R. & Pauly, D. Signature of ocean warming in global fisheries catch. Nature 497, 365–368 (2013).
Tsikliras, A. C., Peristeraki, P., Tserpes, G. & Stergiou, K. I. Mean temperature of the catch (mtc) in the greek seas based on landings and survey data. Front. Marine Sci. 2, 23 (2015).
Leitão, F., Maharaj, R. R., Vieira, V. M., Teodósio, A. & Cheung, W. W. The effect of regional sea surface temperature rise on fisheries along the portuguese iberian atlantic coast. Aquatic Conserv.: Marine Freshw.r Ecosyst. 28, 1351–1359 (2018).
Gianelli, I., Ortega, L., Marin, Y., Piola, A. & Defeo, O. Evidence of ocean warming in uruguay’s fisheries landings: The mean temperature of the catch approach. Marine Ecol. Progr. Series 625, 115–125 (2019).
Kangur, K., Tammiksaar, E. & Pauly, D. Using the “mean temperature of the catch” to assess fish community responses to warming in a temperate lake. Environ. Biol. Fishes (2021).
Pinsky, M. L. & Fogarty, M. Lagged social-ecological responses to climate and range shifts in fisheries. Clim. Change 115, 883–891 (2012).
Rogers, L. A. et al. Shifting habitats expose fishing communities to risk under climate change. Nat. Clim. Change 9, 512–516 (2019).
Selden, R. L. et al. Coupled changes in biomass and distribution drive trends in availability of fish stocks to US West Coast ports. ICES J. Marine Sci. 77, 188–199 (2019).
Nakken, O. Past, present and future exploitation and management of marine resources in the barents sea and adjacent areas. Fish. Res. 37, 23–35 (1998).
Kjesbu, O. S. et al. Synergies between climate and management for atlantic cod fisheries at high latitudes. Proc. Natl. Acad. Sci. 111, 3478–3483 (2014).
Sandø, A. B., Johansen, G. O., Aglen, A., Stiansen, J. E. & Renner, A. H. H. Climate change and new potential spawning sites for northeast arctic cod. Front. Marine Sci. 7, 28 (2020).
Bockstael, N. E. & Opaluch, J. J. Discrete modelling of supply response under uncertainty: The case of the fishery. J. Environ. Econ. Manag. 10, 125–137 (1983).
Holland, D. S. & Sutinen, J. G. Location choice in new england trawl fisheries: Old habits die hard. Land Econ. 76, 133–149 (2000).
Abbott, J. K. & Wilen, J. E. Dissecting the tragedy: A spatial model of behavior in the commons. J. Environ. Econ. Manag. 62, 386–401 (2011).
McQueen, K. & Marshall, C. T. Shifts in spawning phenology of cod linked to rising sea temperatures. ICES J. Marine Sci. 74, 1561–1573 (2017).
Pauly, D. & Liang, C. Temperature and the maturation of fish: a simple sine-wave model for predicting accelerated spring spawning. Environ. Biol. Fishes 105, 1481–1487 (2022).
Sundby, S. & Nakken, O. Spatial shifts in spawning habitats of Arcto-Norwegian cod related to multidecadal climate oscillations and climate change. ICES J. Marine Sci. 65, 953–962 (2008).
Fossheim, M. et al. Recent warming leads to a rapid borealization of fish communities in the arctic. Nat. Clim. Change 5, 673–677 (2015).
Langangen, Ø. et al. Ticket to spawn: Combining economic and genetic data to evaluate the effect of climate and demographic structure on spawning distribution in atlantic cod. Global Change Biol. 25, 134–143 (2019).
Girardin, R. et al. Thirty years of fleet dynamics modelling using discrete-choice models: What have we learned? Fish Fisheries 18, 638–655 (2017).
Wijermans, N., Boonstra, W. J., Orach, K., Hentati-Sundberg, J. & Schlüter, M. Behavioural diversity in fishing—towards a next generation of fishery models. Fish Fisheries 21, 872–890 (2020).
Diekert, F. K. & Nieminen, E. International fisheries agreements with a shifting stock. Dynamic Games Appl. 7, 185–211 (2017).
Young, T. et al. Adaptation strategies of coastal fishing communities as species shift poleward. ICES J. Marine Sci. 76, 93–103 (2018).
Miller, D. D., Ota, Y., Sumaila, U. R., Cisneros-Montemayor, A. M. & Cheung, W. W. L. Adaptation strategies to climate change in marine systems. Global Change Biol. 24, e1–e14 (2018).
Robinson, J. P. W. et al. Diversification insulates fisher catch and revenue in heavily exploited tropical fisheries. Sci. Adv. 6, eaaz0587 (2020).
Kasperski, S. & Holland, D. S. Income diversification and risk for fishermen. Proc. Natl. Acad. Sci. 110, 2076–2081 (2013).
Fernandes, J. A. et al. Estimating the ecological, economic and social impacts of ocean acidification and warming on uk fisheries. Fish Fisheries 18, 389–411 (2017).
Payne, M. R., Kudahl, M., Engelhard, G. H., Peck, M. A. & Pinnegar, J. K. Climate risk to european fisheries and coastal communities. Proc. Natl. Acad. Sci. 118, e2018086118 (2021).
Hannesson, R. Sharing the northeast arctic Cod: Possible effects of climate change. Natl. Resource Model. 19, 633–654 (2006).
Eide, A. An integrated study of economic effects of and vulnerabilities to global warming on the barents sea cod fisheries. Clim. Change 87, 251–262 (2008).
Shchepetkin, A. F. & McWilliams, J. C. The regional oceanic modeling system (roms): a split-explicit, free-surface, topography-following-coordinate oceanic model. Ocean Model. 9, 347–404 (2005).
Haidvogel, D. et al. Ocean forecasting in terrain-following coordinates: Formulation and skill assessment of the regional ocean modeling system. J. Comput. Phys.227, 3595–3624 (2008). Predicting weather, climate and extreme events.
Lien, V. S., Gusdal, Y. & Vikebø, F. B. Along-shelf hydrographic anomalies in the nordic seas (1960–2011): locally generated or advective signals? Ocean Dyn. 64, 1047–1059 (2014).
Lien, V. S., Gusdal, Y., Albretsen, J., Melsom, A. & Vikebø, F. B. Evaluation of a nordic seas 4 km numerical ocean model hindcast archive (svim), 1960-2011. Tech. Rep., Havforskningsinstituttet (2013).
Meager, J. J. et al. Vertical dynamics and reproductive behaviour of farmed and wild atlantic cod gadus morhua. Mar. Ecol. Prog. Ser. 389, 233–243 (2009).
Bergé, L. Efficient estimation of maximum likelihood models with multiple fixed-effects: the R package FENmlm. CREA Discussion Papers (2018).
Svim ocean hindcast archive [data set] (2015).
Acknowledgements
This research has been funded by the Norwegian Research Council Projects FishCom (NFR Nr 280467) and FishTech (NFR Nr 280541).
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
K.A., F.D., and Ø.L. designed research, A.M. provided data and interpretation, K.A. analyzed the data, F.D. and K.A. wrote the first draft of the paper, all authors approved the final draft.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abe, K., Diekert, F., Melsom, A. et al. Do fishers follow fish displaced by climate warming?. npj Ocean Sustain 3, 27 (2024). https://doi.org/10.1038/s44183-024-00063-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s44183-024-00063-9
